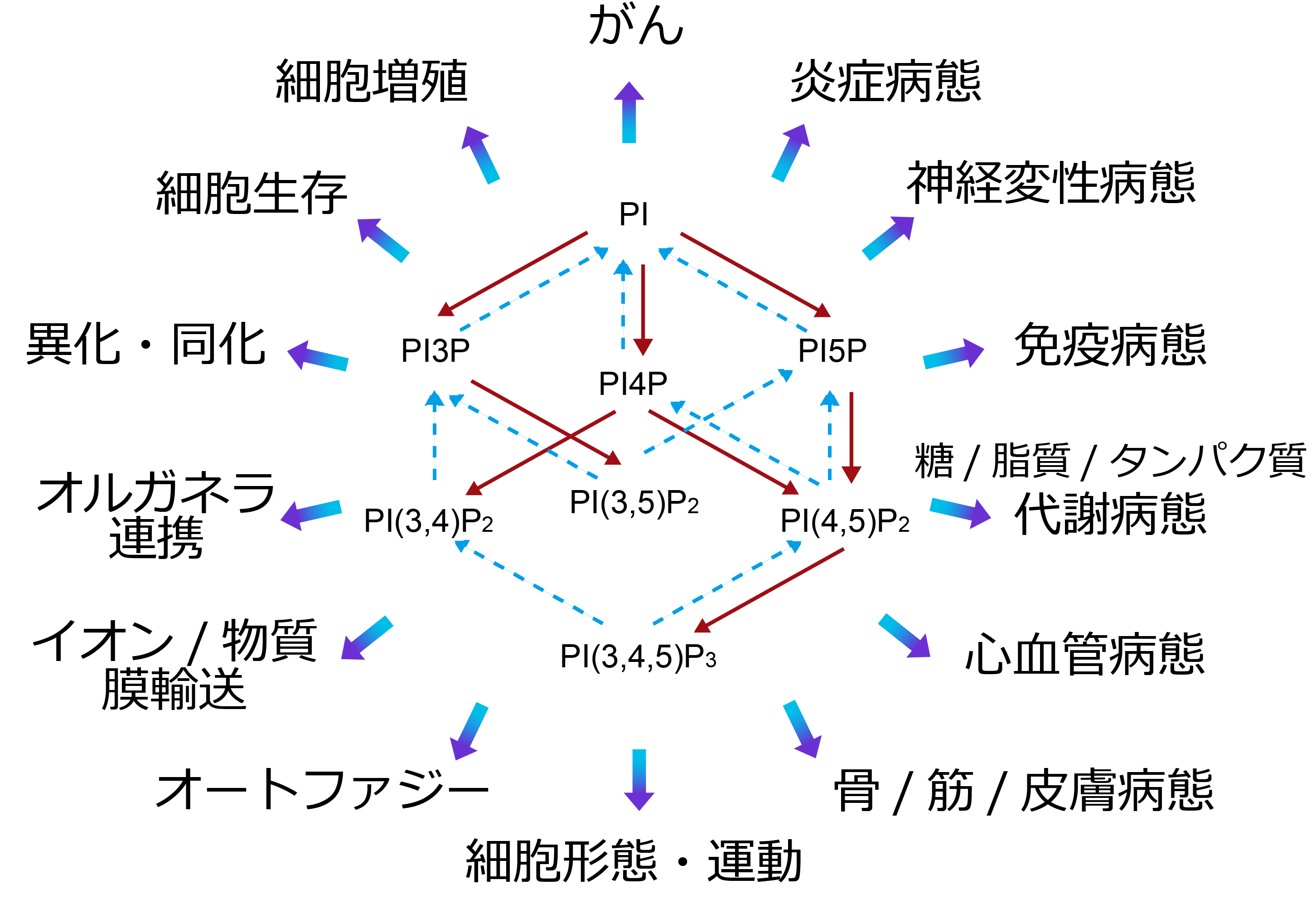
PI accounts for 1–10% of membrane phospholipids. A fraction is phosphorylated to generate the phosphoinositides below. In normal cells and tissues, their abundance is roughly maintained in the order PI ≫ PI4P ≥ PI(4,5)P2 > PI3P > PI(3,5)P2 > PI(3,4,5)P3 ≫ PI(3,4)P2, PI5P.
Present at <1% of PI. It plays an important role in regulating protein degradation. In the formation of the ESCRT complex, which sorts ubiquitinated proteins into multivesicular bodies, it recruits Hrs (ESCRT-0) and EAP45 (ESCRT-II). In autophagy, it targets WIPI2 and DFCP1 to autophagosome-initiation sites and, via Alfy, promotes accumulation of ubiquitinated proteins into autophagosomes. Through binding to SNX5, kinesin-16B, and others, it participates in many aspects of intracellular vesicular transport.
Nat Med 18:1560-1569, 2012
JCI Insight 2:e89462, 2017
Present at 3–5% of PI. Its functions at the Golgi and plasma membrane have attracted attention. It recruits AP1 to the Golgi to promote formation of cargo-laden clathrin-coated vesicles, and via GOLPH3 it controls membrane morphology and vesicle budding. It also plays an important role in lipid transport at contact sites — for example, CERT-mediated ceramide transfer from the ER to the Golgi, ORP-mediated transport of phosphatidylserine from the ER to the plasma membrane, and cholesterol transport from the ER to the Golgi. Although with low specificity, it also acts on plasma-membrane transmembrane proteins, with reported involvement in regulating the TRPV1 cation channel and the mechanosensitive Piezo cation channel.
Nat Commun 16:7808, 2025
J Cell Biol 221:e202103141, 2022
Usually present only in trace amounts, and much about its physiological functions remains unknown. Its nuclear functions have been studied: it binds ING2 via a plant homeodomain (PHD) finger and participates in the DNA-damage response through stabilization of p53, and it binds the basic amino-acid clusters of SAP30 and SAP30L to regulate histone deacetylation. It has also been reported to induce autophagy via PI3P-target proteins under glucose starvation. However, few papers exist, and it remains a lipid full of mysteries.
Usually present only in trace amounts, but produced intracellularly upon receptor stimulation by growth factors and chemotactic factors or by hydrogen-peroxide treatment. In cancer tissue it can accumulate to levels comparable to PI(4,5)P2. By binding several SNX-family molecules, it promotes maturation and scission of clathrin-coated vesicles and macropinosomes. In cell morphology and motility, it recruits lamellipodin (Lpd) to the plasma membrane and, through Ena/VASP-mediated actin-filament elongation and WAVE-mediated actin-filament branching, promotes ruffling and lamellipodia formation. Its binding to p40phox on the phagosome membrane is essential for reactive-oxygen production by NADPH-oxidase assembly. Binding of αTTP to PI(3,4)P2 promotes vitamin E release and is involved in its handoff to ABC transporters at the plasma membrane. In the nucleus, it is involved in oxidative-stress-associated DNA damage.
Nature 465:497-501, 2010
Mol Cell 68:566-580, 2017
J Biochem mvag027, 2026
Present at <0.5% of PI. Its functions are noted mainly in endosomes and lysosomes; depletion of PI(3,5)P2 forms vacuoles that are visible even by light microscopy. Acting on ion channels, it activates TRPML1 to promote Ca2+ release from lysosomes and activates two-pore channels (TPC) 1/2 to promote Na+ release from lysosomes and melanosomes. It binds Vps24 of the ESCRT-III complex and is involved in protein degradation via the multivesicular-body pathway.
Nat Commun 17:4602, 2026
J Cell Biol 224:e202408174, 2025
Present at 3–5% of PI. It serves as a substrate for PLC, producing diacylglycerol and inositol (1,4,5)-trisphosphate. Elucidation of the binding and regulatory functions of PI(4,5)P2 toward actin-polymerization regulators such as profilin and α-actinin became the trigger for broad study of the direct actions of inositol phospholipids. It mediates focal-adhesion formation via the ezrin/radixin/moesin (ERM) proteins that crosslink actin filaments to the plasma membrane, as well as talin and vinculin. The membrane transporters regulated by PI(4,5)P2, including ion channels and transporters, are too numerous to list. Plasma-membrane functions — such as AP-2-mediated clathrin assembly in clathrin-coated-pit formation and epsin-1-mediated membrane deformation — were studied first, but involvement in membrane-protein recycling and autophagy in the endosomal system has also been suggested. Because it activates cPLA2 and PLD, it is thought to participate in other phospholipid metabolism through generation of lysophospholipids and phosphatidic acid.
Nat Commun 16:8852, 2025
Nat Commun 13:2347, 2022
Present at <0.1% of PI, but in cancer tissue it can rise to about 100-fold. Because its degrading enzymes are highly active and because class I phosphoinositide 3-kinase (PI3K) is activated in a receptor-stimulation-dependent manner by various extracellular agonists such as insulin, it functions as a second messenger. Its functions are understood mainly at the plasma membrane. Its binding proteins are diverse, including protein kinases such as PKB and Btk; ion channels such as TRPM3; GEFs for the small GTPases Rac and Arf6 and GAPs for Rho; myosin-X and various actin-binding proteins; and cytoskeleton-related proteins such as WAVE2. It governs cell proliferation, differentiation, migration, cell death, and the regulation of glucose and lipid metabolism.
Cell 95:29-39, 1998
Science 287:1040-1046, 2000
Science 324:384-387, 2009